CONCEPTOS BÁSICOS DE CINÉTICA QUÍMICA
En una reacción de orden cero, la velocidad de formación del producto es independiente de la concentración de sustrato: v = k
En una reacción de primer orden la velocidad de formación de los productos es directamente proporcional a la concentración del sustrato: v = k [A]. Así, en la reacción:
sacarosa + agua
glucosa + fructosa
La velocidad de hidrólisis de la sacarosa es, en todo momento, proporcional a la concentración de sacarosa. Dicho matemáticamente, donde [A] es la concentración de sacarosa a cada tiempo (t) y k es la constante de proporcionalidad. Se dice que ésta es una reacción de primer orden.
Una reacción de segundo orden es aquella en la que la velocidad de formación del producto depende
de la concentración de dos sustratos (como en una reacción de condensación): v = k [A1] [A2]
del cuadrado de la concentración de un único sustrato (reacción de dimerización): v = k [A]2
CINÉTICA ENZIMÁTICA
La cinética enzimática estudia la velocidad de las reacciones catalizadas por enzimas. Estos estudios proporcionan información directa acerca del mecanismo de la reacción catalítica y de la especifidad del enzima. La velocidad de una reacción catalizada por un enzima puede medirse con relativa facilidad, ya que en muchos casos no es necesario purificar o aislar el enzima. La medida se realiza siempre en las condiciones óptimas de pH, temperatura, presencia de cofactores, etc, y se utilizan concentraciones saturantes de sustrato. En estas condiciones, la velocidad de reacción observada es la velocidad máxima (Vmax). La velocidad puede determinarse bien midiendo la aparición de los productos o la desaparición de los reactivos.
Al seguir la velocidad de aparición de producto (o de desaparición del sustrato) en función del tiempo se obtiene la llamada curva de avance de la reacción, o simplemente, la cinética de la reacción. A medida que la reacción transcurre, la velocidad de acumulación del producto va disminuyendo porque se va consumiendo el sustrato de la reacción (Figura de la derecha). Para evitar esta complicación se procede a medir la velocidad inicial de la reacción (v0). La velocidad inicial de la reacción es igual a la pendiente de la curva de avance a tiempo cero (Figura de la derecha). De esta forma, la medida de v0 se realiza antes de que se consuma el 10% del total del sustrato, de forma que pueda considerarse la [S] como esencialmente constante a lo largo del experimento. Además, en estas condiciones no es necesario considerar la reacción inversa, ya que la cantidad de producto formada es tan pequeña que la reacción inversa apenas ocurre. De esta forma se simplifican enormemente las ecuaciones de velocidad.
Para estudiar la cinética enzimática se mide el efecto de la concentración inicial de sustrato sobre la velocidad inicial de la reacción, manteniendo la cantidad de enzima constante. Si representamos v0 frente a [S]0 obtenemos una gráfica como la de la Figura de la derecha. Cuando [S]0 es pequeña, la velocidad inicial es directamente proporcional a la concentración de sustrato, y por tanto, la reacción es de primer orden. A altas [S]0, el enzima se encuentra saturada por el sustrato, y la velocidad ya no depende de [S]0. En este punto, la reacción es de orden cero y la velocidad es máxima (Vmax).
lunes, 11 de mayo de 2009
***bIoEnerGétiCa***
La Bioenergética es la parte de la biología muy relacionada con la física, que se encarga del estudio de los procesos de absorción transformación y entrega de energía en los sistemas biológicos. En general, la Bioenergética se relaciona con la Termodinámica, en particular con el tema de la Energía Libre, en especial la Energía Libre de Gibbs. Los cambios en la energía libre de Gibbs ΔG nos dan una cuantificación de la factibilidad energética de una reacción química y pueden proveer de una predicción de si la reacción podrá suceder o no. Como una característica general de La Bioenergética, esta solo se interesa por los estados energéticos inicial y final de los componentes de una reacción química, los tiempos necesarios para que el cambio químico se lleve a cabo en general se desprecian. Un objetivo general de la Bioenergética, es predecir si ciertos procesos son posibles o no; en general, la cinética cuantifica qué tan rápido ocurre la reacción química.
El metabolismo es el conjunto de transformaciones que experimenta la materia externa desde su absorción o adición al citoplasma, hasta su eliminación del mismo. Por ejemplo, las células están compuestas por un complejo sistema de reacciones químicas que generan energía y otras que utilizan energía, esto en general es el Metabolismo. El Metabolismo Comprende dos fases:
El anabolismo (Síntesis de compuestos orgánicos)
El catabolismo (Degradación de sustancias complejas)
Estos representan la suma de cambios químicos que convierten los alimentos en formas utilizables de energía y en moléculas biológicas complejas.
En general, el ATP es la conexión entre los sistemas que producen la energía y los que la utilizan; la degradación oxidativa de los alimentos es un proceso exergónico 'son endergónicos y utilizan la energía química almacenada en forma de ATP y NADPH.
Las células vivas son capaces de realizar la conversión de distintas formas de energía y pueden intercambiar energía con su entorno, es conveniente revisar algunas leyes o principios de la termodinámica que rigen las reacciones de este tipo. El primer principio de la termodinámica es una ley de conservación de la energía y estipula que, aunque la energía se puede convertir de una forma a otra, la energía total del sistema ha de permanecer constante. Por ejemplo, la energía química disponible en un combustible metabólico tal como la glucosa se puede convertir en el proceso de la glucólisis en otra forma de energía química, el ATP. La energía implicada en un gradiente osmótico electro potencial de protones establecido a través de la membrana mitocondrial puede convertirse en energía química al utilizar dicho gradiente para impulsar la síntesis de ATP. Para discutir el segundo principio de la Termodinámica se debe definir el término entropía. La entropía (que se designa con el símbolo S) es una medida o indicador del grado de desorden en un sistema. La entropía se puede considerar también como la energía de un sistema que no se puede utilizar para realizar trabajo efectivo. Todos los procesos, ya sean químicos o biológicos progresan hacia una situación de máxima entropía. No obstante, en los sistemas biológicos es casi imposible cuantificar cambios de entropía ya que estos sistemas raramente están en equilibrio. Por razones de sencillez y por su utilidad inherente en estos tipos de consideraciones, se empleará la cantidad denominada energía libre.
El metabolismo es el conjunto de transformaciones que experimenta la materia externa desde su absorción o adición al citoplasma, hasta su eliminación del mismo. Por ejemplo, las células están compuestas por un complejo sistema de reacciones químicas que generan energía y otras que utilizan energía, esto en general es el Metabolismo. El Metabolismo Comprende dos fases:
El anabolismo (Síntesis de compuestos orgánicos)
El catabolismo (Degradación de sustancias complejas)
Estos representan la suma de cambios químicos que convierten los alimentos en formas utilizables de energía y en moléculas biológicas complejas.
En general, el ATP es la conexión entre los sistemas que producen la energía y los que la utilizan; la degradación oxidativa de los alimentos es un proceso exergónico 'son endergónicos y utilizan la energía química almacenada en forma de ATP y NADPH.
Las células vivas son capaces de realizar la conversión de distintas formas de energía y pueden intercambiar energía con su entorno, es conveniente revisar algunas leyes o principios de la termodinámica que rigen las reacciones de este tipo. El primer principio de la termodinámica es una ley de conservación de la energía y estipula que, aunque la energía se puede convertir de una forma a otra, la energía total del sistema ha de permanecer constante. Por ejemplo, la energía química disponible en un combustible metabólico tal como la glucosa se puede convertir en el proceso de la glucólisis en otra forma de energía química, el ATP. La energía implicada en un gradiente osmótico electro potencial de protones establecido a través de la membrana mitocondrial puede convertirse en energía química al utilizar dicho gradiente para impulsar la síntesis de ATP. Para discutir el segundo principio de la Termodinámica se debe definir el término entropía. La entropía (que se designa con el símbolo S) es una medida o indicador del grado de desorden en un sistema. La entropía se puede considerar también como la energía de un sistema que no se puede utilizar para realizar trabajo efectivo. Todos los procesos, ya sean químicos o biológicos progresan hacia una situación de máxima entropía. No obstante, en los sistemas biológicos es casi imposible cuantificar cambios de entropía ya que estos sistemas raramente están en equilibrio. Por razones de sencillez y por su utilidad inherente en estos tipos de consideraciones, se empleará la cantidad denominada energía libre.
sábado, 9 de mayo de 2009
°°°°pRopIeDadEs dE laS prOteInaS°°°
Son 3 las propiedades que tienen las proteínas desde el punto de vista bioquímico:
- a) precipitación selectiva
- b) capacidad amortiguadora
- c) propiedades osmóticas
PRECIPITACIÓ N SELECTIVA DE LAS PROTEÍNAS
N SELECTIVA DE LAS PROTEÍNAS
El agua es el disolvente biológico por excelencia. En disolución acuosa, los residuos hidrofóbicos de las proteínas se acumulan en el interior de la estructura, mientras que en la superficie aparecen diversos grupos con carga eléctrica, en función del pH del medio. En torno a los grupos cargados, los dipolos del agua se orientan conforme a la carga eléctrica de cada grupo, de tal manera que la proteína presenta una capa de solvatación formada por el agua de hidratación, que es el agua retenida por las cargas eléctricas de la superficie de las proteínas. Los AA polares sin carga también se disponen en la superficie, donde interaccionan con el agua mediante puentes de hidrógeno.
Cualquier factor que modifique la interacción de la proteína con el disolvente disminuirá su estabilidad en disolución y provocará la precipitación. Así, la desaparición total o parcial de la envoltura acuosa, la neutralización de las cargas eléctricas de tipo repulsivo o la ruptura de los puentes de hidrógeno facilitará la agregación intermolecular y provocará la precipitación. La precipitación suele ser consecuencia del fenómeno llamado desnaturalización.
Capacidad amortiguadora de las proteínas
Esta propiedad se debe a la existencia de:
Grupos ionizables de las cadenas laterales de los aminoácidos Asp, Glu, Lys, Arg, His, Tyr, Cys.
Grupos COOH y NH2 terminales.
Por este motivo, las proteínas poseen un considerable poder amortiguador en una amplia zona de pH. Aunque cada AA tiene unos grupos ionizables con unas constantes de ionización (pKa) características, el valor de dichas constantes puede verse ligeramente modificado por el entorno proteico. El grupo imidazol del AA histidina es el principal responsable del poder amortiguador de las proteínas a pH fisiológico, ya que su pKa está próximo a 7.
Cuando el pH es bajo, los grupos ionizables están protonados, y la carga neta de la proteína es de signo positivo. Cuando el pH es alto, los grupos ionizables están desprotonados, y la carga neta es de signo negativo. Entre ambas zonas, habrá un pH en el cual la carga neta de la proteína es nula. Es el pH isoeléctrico o punto isoeléctrico, y es característico de cada proteína.
A valores de pH por debajo del pH isoeléctrico la carga neta de la proteína es positiva, y a valores de pH por encima del pH isoeléctrico, la carga neta de la proteína es negativa. La mayoría de las proteínas intracelulares tienen carga negativa, ya que su pH isoeléctrico es menor que el pH fisiológico (que está proximo a 7). Se llaman proteínas ácidas a aquellas que tienen un punto isoeléctrico bajo (como la pepsina), y proteínas básicas a las que tienen un punto isoeléctrico alto (como las histonas).
 N SELECTIVA DE LAS PROTEÍNAS
N SELECTIVA DE LAS PROTEÍNAS El agua es el disolvente biológico por excelencia. En disolución acuosa, los residuos hidrofóbicos de las proteínas se acumulan en el interior de la estructura, mientras que en la superficie aparecen diversos grupos con carga eléctrica, en función del pH del medio. En torno a los grupos cargados, los dipolos del agua se orientan conforme a la carga eléctrica de cada grupo, de tal manera que la proteína presenta una capa de solvatación formada por el agua de hidratación, que es el agua retenida por las cargas eléctricas de la superficie de las proteínas. Los AA polares sin carga también se disponen en la superficie, donde interaccionan con el agua mediante puentes de hidrógeno.
Cualquier factor que modifique la interacción de la proteína con el disolvente disminuirá su estabilidad en disolución y provocará la precipitación. Así, la desaparición total o parcial de la envoltura acuosa, la neutralización de las cargas eléctricas de tipo repulsivo o la ruptura de los puentes de hidrógeno facilitará la agregación intermolecular y provocará la precipitación. La precipitación suele ser consecuencia del fenómeno llamado desnaturalización.
Capacidad amortiguadora de las proteínas
Esta propiedad se debe a la existencia de:
Grupos ionizables de las cadenas laterales de los aminoácidos Asp, Glu, Lys, Arg, His, Tyr, Cys.
Grupos COOH y NH2 terminales.
Por este motivo, las proteínas poseen un considerable poder amortiguador en una amplia zona de pH. Aunque cada AA tiene unos grupos ionizables con unas constantes de ionización (pKa) características, el valor de dichas constantes puede verse ligeramente modificado por el entorno proteico. El grupo imidazol del AA histidina es el principal responsable del poder amortiguador de las proteínas a pH fisiológico, ya que su pKa está próximo a 7.
Cuando el pH es bajo, los grupos ionizables están protonados, y la carga neta de la proteína es de signo positivo. Cuando el pH es alto, los grupos ionizables están desprotonados, y la carga neta es de signo negativo. Entre ambas zonas, habrá un pH en el cual la carga neta de la proteína es nula. Es el pH isoeléctrico o punto isoeléctrico, y es característico de cada proteína.
A valores de pH por debajo del pH isoeléctrico la carga neta de la proteína es positiva, y a valores de pH por encima del pH isoeléctrico, la carga neta de la proteína es negativa. La mayoría de las proteínas intracelulares tienen carga negativa, ya que su pH isoeléctrico es menor que el pH fisiológico (que está proximo a 7). Se llaman proteínas ácidas a aquellas que tienen un punto isoeléctrico bajo (como la pepsina), y proteínas básicas a las que tienen un punto isoeléctrico alto (como las histonas).
PROPIEDADES OSM ÓTICAS DE LAS PROTEÍNAS
ÓTICAS DE LAS PROTEÍNAS
Como todo soluto molecular o iónico, las proteínas ejercen un efecto osmótico cuando existen barreras que limitan su libre difusión, como puede ser una membrana semipermeable, que permite el paso del agua, pero no de los solutos. Si tenemos dos compartimentos acuosos separados por una membrana semipermeable y uno de ellos contiene proteínas, éstas tienden a captar agua del compartimento vecino. Este efecto osmótico es proporcional al número de partículas dispersas. El valor de la presión osmótica se puede calcular mediante la fórmula de Van't Hoff: p = cRT, donde p es la
presión osmótica, c es la concentración, R es la constante de los gases y T es la temperatura absoluta.
En el caso de las proteínas, el efecto osmótico se ve amplificado por otros dos factores.
Por un lado, el agua de hidratación que forma la envoltura acuosa de las proteínas también contribuye a la presión osmótica.
Por otro lado, las proteínas se comportan como polianiones, cuyas cargas están neutralizadas por iones Na+ o K+. Las membranas biológicas son permeables a estos iones y a sus contraiones, con lo cual su concentración a ambos lados de la membrana se equilibra. Sin embargo, la existencia de proteínas en sólo uno de los compartimentos provoca la retención permanente de iones difusibles en ese lado de la membrana (efecto Donnan), lo que incrementa el efecto osmótico.
EfectoDonnan: Situación inicial
Efecto Don nan: En el equilibrio
nan: En el equilibrio
Se denomina presión coloidosmótica o presión oncótica al efecto osmótico conjunto de las proteínas, que es el resultado de:
1- la presión osmótica (que sólo depende del número de partículas)
2- la presión provocada por el agua de hidratación
3- la presión provocada por el exceso de iones debido al efecto Donnan
La mayor parte del agua en el sistema circulatorio está retenida por el efecto osmótico de las proteínas del plasma. Cuando por cualquier circunstancia patológica disminuye la concentración de proteínas en el plasma, el agua puede fluir libremente hacia los tejidos, provocando un edema.
iNmunOgLobUliNas!!
ESTRUCTURA DE LAS INMUNOGLOBULINAS
Las inmunoglobulinas son glicoproteínas que, según ya indicó Porter en 1959, están formadas por cadenas polipeptídicas agrupadas, dependiendo del tipo de inmunoglobulina, en una o varias unidades estructurales básicas.
Unidad estructural básica
Cada unidad está compuesta por cuatro cadenas polipeptídicas unidas entre sí por puentes disulfuro y otras uniones de tipo no covalente. Para su estudio se han empleado diferentes procedimientos. Por ejemplo, tras la rotura de los puentes disulfuro por sustancias de carácter reductor, como el mercaptoetanol, se individualizan las cuatro cadenas polipeptídicas y éstos atendiendo a su tamaño, son de dos tipos: de bajo peso molecular (aproximadadamente 22 KD) y de alto peso molecular (50-70 KD, dependiendo del tipo de Ig). Los polipéptidos de bajo peso molecular reciben el nombre de cadenas ligeras o cadenas L (Light) y las de alto peso molecular, cadenas pesadas o cadenas H (Heavy).
Dos cadenas ligeras y dos cadenas pesadas se agrupan de tal manera que existe una proximidad espacial entre los cuatro extremos amínicos de las cadenas ligeras y pesadas por una parte, y entre los dos extremos carboxílicos de las cadenas pesadas por otra.
Esta estructura básica de las inmunoglobulinas puede ser fraccionada mediante la utilización de enzimas (papaína, pepsina, etc.), como fue efectuado por Porter en 1959, obteniéndose diferentes tipos de fragmentos. El tratamiento con papaína produce la ruptura específica de las cadenas H, en el espacio comprendido entre el puente disulfuro que las une entre sí y los que las unen a las cadenas ligeras. Se obtienen tres fragmentos: uno denominado Fc, que determina la actividad biológica, contiene el alotipo y determina la clase y subclase de cadena pesada y dos denominados cada uno de ellos Fab, que contienen el idiotipo y es por donde la molécula se une al antígeno.
Cadenas Ligeras.
Hay dos tipos de cadenas ligeras, estructuralmente diferentes, que se conocen como cadenas ligeras tipo kappa (k) y cadenas ligeras tipo lambda (l). La familia de genes que codifica para la cadena ligera k se localiza en el cromosoma 2 y los loci de los genes homólogos que codifican para la cadena l, en el cromosoma 22. En cada molécula de inmunoglobulina las dos cadenas ligeras son del mismo tipo, k o bien l, pero nunca existe una de cada tipo en la misma inmunoglobulina.
Las cadenas ligeras están formadas por unos 200 aminoácidos con la particularidad de que existen dos puentes disulfuro que unen grupos de unos cincuenta aminoácidos. Concretamente la IgG1 posee 214 aminoácidos y su estructura secundaria y terciaria están determinadas por dos puentes disulfuro intracatenarios que unen los aminoácidos 23 con el 88 y 134 con el 193 . A su vez, estas cadenas ligeras tienen otro puente disulfuro intercatenario, por el cual cada una de ellas se une a una cadena pesada para constituir la unidad básica de las inmunoglobulinas. Este puente se encuentra en el último aminoácido (214) de la parte carboxílica para el tipo k y en el penúltimo para el tipo l.
Cadenas pesadas.
Estas cadenas poseen unos cuatrocientos aminoácidos estableciéndose entre algunos de ellos puentes disulfuro (intracatenarios) que asocian unos 60 aminoácidos y que condicionan la estructura secundaria del polipéptido. Por ejemplo, las cadenas pesadas de la IgG1 poseen 440 aminoácidos y los puentes disulfuro unen el aminoácido 22 con el 96, el 144 con el 200, el 261 con el 321 y el 367 con el 425.
Estas dos cadenas pesadas están unidas la una a la otra por puentes disulfuro intercatenarios, ya indicados anteriormente, y que pueden ser de uno a cinco dependiendo del tipo de inmunoglobulina.
En estas cadenas pesadas, y a nivel de los puentes disulfuro intercatenarios, hay una zona de unos 15 aminoácidos, de gran flexibilidad debido a su estructura y constituye lo que se denomina zona bisagra por donde se deforma la molécula de inmunoglobulina cuando se produce la unión con el antígeno, facilitándose así su acoplamiento con éste. Los loci de los genes que codifican para la cadena pesada se localizan en el brazo largo del cromosoma 14.
FUNCIÓN DE LAS INMUNOGLOBULINAS.
La función esencial de las inmunoglobulinas es la de unirse al antígeno. De esta manera las inmunoglobulinas actúan como receptoras de señales antigénicas o bien pueden colaborar en la destrucción antigénica. La primera función se presenta cuando las inmunoglobulinas se encuentran insertas en la membrana de los linfocitos B (inmunoglobulinas de membrana), y para la segunda requieren la colaboración del complemento, macrófagos, neutrófilos y células NK, que tienen la propiedad de unir las inmunoglobulinas por su extremo Fc.
Unión antígeno anticuerpo.
Los epítopos de un antígeno pueden estar formados por aminoácidos consecutivos en la secuencia de la proteína, como las proteínas se encuentran normalmente dobladas sobre si mismas según lo que llamamos estructura terciaria, en la mayoría de los casos los anticuerpos generados contra este tipo de epítopos solo reconocerán a la proteína desnaturalizada o “linearizada” y por ello se les llama epitopos lineales. En la mayoría de los casos los epítopos suelen estar formados por aminoácidos del antigeno que solo se encuentran suficientemente cerca unos de otros en la proteína nativa, es decir en la proteína que tiene estructura terciaria conservada, es decir una conformación adecuada, por lo que a estos epítopos se les llama epitopos conformacionales. Cuando inmunizamos un animal con una proteína, generaremos una serie de anticuerpos dirigidos contra los distintos epítopos de la misma, todos esos anticuerpos se encontraran circulando en el suero del animal al que, una vez extraido, llamaremos antisuero. El tipo de anticuerpos que compondrán ese antisuero dependerá en gran medida de la forma en que hayamos preparado la proteína para la inmunización, si la hemos preparado desnaturalizada, solo existirán epítopos lineales, mientras que si hemos inyectado la proteína en su estado nativo, coexistirán en el antisuero anticuerpos que reconozcan epítopos conformacionales con otros que reconozcan epítopos lineales. En el caso de anticuerpos monoclonales, todas los anticuerpos procederán de un clon de células plasmáticas y por tanto estarán dirigidos contra un solo epítopo que será de un tipo u otro. La importancia radica, en que dependiendo del tipo de epitopos que reconozcan los anticuerpos, las aplicaciones diagnosticas o de investigación serán distintas. En general, los anticuerpos que reconocen epitopos lineales serán útiles para técnicas de Western Blot (donde se analiza la proteína generalmente desnaturalizada) mientras los que reconocen epítopos conformacionales lo serán para técnicas de inmunofluoresencia, inmunoprecipitación, etc.
Propiedades y función de cada una de las inmunoglobulinas
Aunque en los apartados anteriores se ha hecho mención a las propiedades y función de las inmunoglobulinas, a continuación estudiaremos brevemente y por separado las características funcionales más importantes de cada una de ellas.
Inmunoglobulina G.
Son las inmunoglobulinas más abundantes y representan más del 70 % de las Igs séricas totales; las diferentes subclases se presentan en proporciones muy diferentes. La IgG1 es la subclase más frecuente (más del 60 %), seguida de la IgG2 (aproximadamente un 18 %), mientras que IgG3 e IgG4 se encuentran en mucha menor proporción.
Esta Ig posee capacidad neutralizante, precipitante, de fijar complemento, de unirse a células NK y a macrófagos (opsonización) y son capaces de atravesar activamente las membranas biológicas. La propiedad de atravesar activamente las membranas biológicas es de sumo interés por lo que, además de ejercer esta inmunoglobulina, su efecto en toda la “economía del organismo”, lo hace también en el feto al atravesar la placenta desde la madre, merced a la existencia de receptores para la porción Fc en el sincitiotrofoblasto.
Como el feto sólo sintetiza pequeñas cantidades de inmunoglobulinas, adquiere de este modo la posibilidad de defensa, no solamente mientras se encuentra en el seno materno, sino incluso durante la lactancia, período en el cual todavía no ha desarrollado la capacidad total de síntesis de inmunoglobulinas.
Sin embargo, este paso de IgG desde la madre al feto no siempre es beneficioso para el feto. De todos es sabido que cuando hay incompatibilidad del tipo Rh entre la madre y el feto, se puede desarrollar el síndrome de eritroblastosis fetal como consecuencia de la destrucción de glóbulos rojos fetales, de nefastas consecuencias si no se acude a tiempo. Esto no se presentaría si la IgG no pasase de la madre al feto
La IgG se sintetiza tardíamente tras un primer contacto con el antígeno, sin embargo, tras un segundo contacto la mayoría de las Igs formadas pertenecen a esta clase (Respuesta Secundaria)
Inmunoglobulina M.
Los anticuerpos del tipo IgM son los que mas rápidamente se forman en respuesta a un estímulo antigénico (Respuesta primaria). Esta Ig se caracteriza también por poseer capacidad neutralizante, precipitante, aglutinante, fijar complemento, activar la respuesta inmune, sin embargo no atraviesa activamente las membranas biológicas. Esta última propiedad hace que esta inmunoglobulina ejerza su acción normalmente en los espacios intravasculares.
Representa del 5 al 10 % de las Igs séricas totales y junto a la IgD es la más frecuentemente encontrada en la superficie de los linfocitos B como inmunoglobulina de membrana.
Inmunoglobulina A.
Esta inmunoglobulina posee capacidad neutralizante y precipitante, mientras que su capacidad de fijar complemento y de opsonización son muy débiles, limitándose su efecto a neutrófilos y no a macrófagos.
La propiedad más importante de esta inmunoglobulina viene determinada por su capacidad de unirse por el extremo Fc a la pieza secretora, gracias a la cual puede ser secretada por las mucosas y glándulas exocrinas, ejerciendo su acción más importante en la superficie de mucosas y líquidos biológicos (sobre todo IgA2), tales como el liquido cefaloraquideo, secreción bronquial, lágrima, saliva, etc. Esto es importante porque así protegen precisamente los puntos más vulnerables del organismo, esto es, las puertas de entrada al mismo, como son ojos, boca, aparato digestivo, sistema respiratorio, vagina, etc. No olvidemos que, por ejemplo, si desplegamos la mucosa del aparato respiratorio, la superficie que cubriríamos es de unos 300 m2, superficie que se encuentra en contacto directo con el exterior a través del aire que se respira. Se deduce de ello que, sin duda, deben ser importantes los mecanismos de defensa local entre los cuales la IgA tiene un papel esencial.
Esta inmunoglobulina se encuentra también en la leche materna. Los niveles de todas las inmunoglobulinas, a excepción de la IgG en recién nacidos son muy bajos, siendo por tanto de gran significación el hecho de que la IgA se transfiera desde la madre al lactante a través de la secreción láctea. De ahí que tengamos que insistir en que los lactantes se amamanten en el mayor grado posible directamente por las madres y no con leche de otros orígenes, a lo que actualmente existe excesiva tendencia.
La IgA recibida de la madre ejerce un importante papel de defensa a nivel de todo el aparato digestivo. En ello parece que influyen las especiales características de pH gástrico del lactante que es menos ácido que en el adulto y una especial resistencia de esta inmunoglobulina frente al mismo, por lo que no se destruye a su paso por el estómago.
Inmunoglobulina D.
La concentración de esta inmunoglobulina en suero es muy baja. Hasta fechas muy recientes no se había demostrado que esta inmunoglobulina poseía capacidad de unirse a antígenos, por lo que se dudaba de que actuase con función de anticuerpo. Sin embargo, aunque actualmente se ha demostrado su acción de anticuerpo, no se conoce con precisión cuáles son sus funciones específicas, aunque se piensa que colabora de forma importante en la activación de linfocitos B al actuar como receptor en la superficie de los mismos.
Inmunoglobulina E.
En muchos individuos alérgicos esta inmunoglobulina se presenta en grandes cantidades. El estímulo para su síntesis puede proceder de una gran variedad de antígenos, a los que en este caso se conocen como alergenos. Estos alergenos pueden penetrar en el organismo a través de la piel o de las mucosas respiratoria, ocular, del aparato digestivo, etc., así como por inyectables, como es el caso de la penicilina u otros medicamentos.
La vida media de la IgE en sangre periférica es de 24-48 horas. No tiene capacidad de atravesar la placenta, por lo tanto, las reacciones de hipersensibilidad inmediata no pueden transferirse de manera pasiva de la madre al feto. Sin embargo, puede existir una predisposición de tipo familiar a padecer enfermedades de naturaleza alérgica. Esta predisposición parece estar relacionada con una tendencia a producir anticuerpos de tipo IgE en la respuesta secundaria frente a antígenos, en lugar de IgG que seria la respuesta normal en individuos no alérgicos.
La IgE se encuentra en forma libre en sangre, tambien en otros líquidos biológicos así como unida a basófilos y células cebadas, gracias a la propiedad que tiene esta inmunoglobulina de unirse por su extremo Fc a receptores de superficie presentes en dichas células. Estas células se caracterizan por encontrarse en la piel y mucosas y por contener abundantes gránulos citoplasmáticos, ricos en sustancias vasoactivas que liberan una vez se activan.
Las inmunoglobulinas son glicoproteínas que, según ya indicó Porter en 1959, están formadas por cadenas polipeptídicas agrupadas, dependiendo del tipo de inmunoglobulina, en una o varias unidades estructurales básicas.
Unidad estructural básica
Cada unidad está compuesta por cuatro cadenas polipeptídicas unidas entre sí por puentes disulfuro y otras uniones de tipo no covalente. Para su estudio se han empleado diferentes procedimientos. Por ejemplo, tras la rotura de los puentes disulfuro por sustancias de carácter reductor, como el mercaptoetanol, se individualizan las cuatro cadenas polipeptídicas y éstos atendiendo a su tamaño, son de dos tipos: de bajo peso molecular (aproximadadamente 22 KD) y de alto peso molecular (50-70 KD, dependiendo del tipo de Ig). Los polipéptidos de bajo peso molecular reciben el nombre de cadenas ligeras o cadenas L (Light) y las de alto peso molecular, cadenas pesadas o cadenas H (Heavy).
Dos cadenas ligeras y dos cadenas pesadas se agrupan de tal manera que existe una proximidad espacial entre los cuatro extremos amínicos de las cadenas ligeras y pesadas por una parte, y entre los dos extremos carboxílicos de las cadenas pesadas por otra.
Esta estructura básica de las inmunoglobulinas puede ser fraccionada mediante la utilización de enzimas (papaína, pepsina, etc.), como fue efectuado por Porter en 1959, obteniéndose diferentes tipos de fragmentos. El tratamiento con papaína produce la ruptura específica de las cadenas H, en el espacio comprendido entre el puente disulfuro que las une entre sí y los que las unen a las cadenas ligeras. Se obtienen tres fragmentos: uno denominado Fc, que determina la actividad biológica, contiene el alotipo y determina la clase y subclase de cadena pesada y dos denominados cada uno de ellos Fab, que contienen el idiotipo y es por donde la molécula se une al antígeno.

Cadenas Ligeras.
Hay dos tipos de cadenas ligeras, estructuralmente diferentes, que se conocen como cadenas ligeras tipo kappa (k) y cadenas ligeras tipo lambda (l). La familia de genes que codifica para la cadena ligera k se localiza en el cromosoma 2 y los loci de los genes homólogos que codifican para la cadena l, en el cromosoma 22. En cada molécula de inmunoglobulina las dos cadenas ligeras son del mismo tipo, k o bien l, pero nunca existe una de cada tipo en la misma inmunoglobulina.
Las cadenas ligeras están formadas por unos 200 aminoácidos con la particularidad de que existen dos puentes disulfuro que unen grupos de unos cincuenta aminoácidos. Concretamente la IgG1 posee 214 aminoácidos y su estructura secundaria y terciaria están determinadas por dos puentes disulfuro intracatenarios que unen los aminoácidos 23 con el 88 y 134 con el 193 . A su vez, estas cadenas ligeras tienen otro puente disulfuro intercatenario, por el cual cada una de ellas se une a una cadena pesada para constituir la unidad básica de las inmunoglobulinas. Este puente se encuentra en el último aminoácido (214) de la parte carboxílica para el tipo k y en el penúltimo para el tipo l.
Cadenas pesadas.
Estas cadenas poseen unos cuatrocientos aminoácidos estableciéndose entre algunos de ellos puentes disulfuro (intracatenarios) que asocian unos 60 aminoácidos y que condicionan la estructura secundaria del polipéptido. Por ejemplo, las cadenas pesadas de la IgG1 poseen 440 aminoácidos y los puentes disulfuro unen el aminoácido 22 con el 96, el 144 con el 200, el 261 con el 321 y el 367 con el 425.
Estas dos cadenas pesadas están unidas la una a la otra por puentes disulfuro intercatenarios, ya indicados anteriormente, y que pueden ser de uno a cinco dependiendo del tipo de inmunoglobulina.
En estas cadenas pesadas, y a nivel de los puentes disulfuro intercatenarios, hay una zona de unos 15 aminoácidos, de gran flexibilidad debido a su estructura y constituye lo que se denomina zona bisagra por donde se deforma la molécula de inmunoglobulina cuando se produce la unión con el antígeno, facilitándose así su acoplamiento con éste. Los loci de los genes que codifican para la cadena pesada se localizan en el brazo largo del cromosoma 14.
FUNCIÓN DE LAS INMUNOGLOBULINAS.
La función esencial de las inmunoglobulinas es la de unirse al antígeno. De esta manera las inmunoglobulinas actúan como receptoras de señales antigénicas o bien pueden colaborar en la destrucción antigénica. La primera función se presenta cuando las inmunoglobulinas se encuentran insertas en la membrana de los linfocitos B (inmunoglobulinas de membrana), y para la segunda requieren la colaboración del complemento, macrófagos, neutrófilos y células NK, que tienen la propiedad de unir las inmunoglobulinas por su extremo Fc.
Unión antígeno anticuerpo.
Los epítopos de un antígeno pueden estar formados por aminoácidos consecutivos en la secuencia de la proteína, como las proteínas se encuentran normalmente dobladas sobre si mismas según lo que llamamos estructura terciaria, en la mayoría de los casos los anticuerpos generados contra este tipo de epítopos solo reconocerán a la proteína desnaturalizada o “linearizada” y por ello se les llama epitopos lineales. En la mayoría de los casos los epítopos suelen estar formados por aminoácidos del antigeno que solo se encuentran suficientemente cerca unos de otros en la proteína nativa, es decir en la proteína que tiene estructura terciaria conservada, es decir una conformación adecuada, por lo que a estos epítopos se les llama epitopos conformacionales. Cuando inmunizamos un animal con una proteína, generaremos una serie de anticuerpos dirigidos contra los distintos epítopos de la misma, todos esos anticuerpos se encontraran circulando en el suero del animal al que, una vez extraido, llamaremos antisuero. El tipo de anticuerpos que compondrán ese antisuero dependerá en gran medida de la forma en que hayamos preparado la proteína para la inmunización, si la hemos preparado desnaturalizada, solo existirán epítopos lineales, mientras que si hemos inyectado la proteína en su estado nativo, coexistirán en el antisuero anticuerpos que reconozcan epítopos conformacionales con otros que reconozcan epítopos lineales. En el caso de anticuerpos monoclonales, todas los anticuerpos procederán de un clon de células plasmáticas y por tanto estarán dirigidos contra un solo epítopo que será de un tipo u otro. La importancia radica, en que dependiendo del tipo de epitopos que reconozcan los anticuerpos, las aplicaciones diagnosticas o de investigación serán distintas. En general, los anticuerpos que reconocen epitopos lineales serán útiles para técnicas de Western Blot (donde se analiza la proteína generalmente desnaturalizada) mientras los que reconocen epítopos conformacionales lo serán para técnicas de inmunofluoresencia, inmunoprecipitación, etc.
Propiedades y función de cada una de las inmunoglobulinas
Aunque en los apartados anteriores se ha hecho mención a las propiedades y función de las inmunoglobulinas, a continuación estudiaremos brevemente y por separado las características funcionales más importantes de cada una de ellas.
Inmunoglobulina G.
Son las inmunoglobulinas más abundantes y representan más del 70 % de las Igs séricas totales; las diferentes subclases se presentan en proporciones muy diferentes. La IgG1 es la subclase más frecuente (más del 60 %), seguida de la IgG2 (aproximadamente un 18 %), mientras que IgG3 e IgG4 se encuentran en mucha menor proporción.
Esta Ig posee capacidad neutralizante, precipitante, de fijar complemento, de unirse a células NK y a macrófagos (opsonización) y son capaces de atravesar activamente las membranas biológicas. La propiedad de atravesar activamente las membranas biológicas es de sumo interés por lo que, además de ejercer esta inmunoglobulina, su efecto en toda la “economía del organismo”, lo hace también en el feto al atravesar la placenta desde la madre, merced a la existencia de receptores para la porción Fc en el sincitiotrofoblasto.
Como el feto sólo sintetiza pequeñas cantidades de inmunoglobulinas, adquiere de este modo la posibilidad de defensa, no solamente mientras se encuentra en el seno materno, sino incluso durante la lactancia, período en el cual todavía no ha desarrollado la capacidad total de síntesis de inmunoglobulinas.
Sin embargo, este paso de IgG desde la madre al feto no siempre es beneficioso para el feto. De todos es sabido que cuando hay incompatibilidad del tipo Rh entre la madre y el feto, se puede desarrollar el síndrome de eritroblastosis fetal como consecuencia de la destrucción de glóbulos rojos fetales, de nefastas consecuencias si no se acude a tiempo. Esto no se presentaría si la IgG no pasase de la madre al feto
La IgG se sintetiza tardíamente tras un primer contacto con el antígeno, sin embargo, tras un segundo contacto la mayoría de las Igs formadas pertenecen a esta clase (Respuesta Secundaria)
Inmunoglobulina M.
Los anticuerpos del tipo IgM son los que mas rápidamente se forman en respuesta a un estímulo antigénico (Respuesta primaria). Esta Ig se caracteriza también por poseer capacidad neutralizante, precipitante, aglutinante, fijar complemento, activar la respuesta inmune, sin embargo no atraviesa activamente las membranas biológicas. Esta última propiedad hace que esta inmunoglobulina ejerza su acción normalmente en los espacios intravasculares.
Representa del 5 al 10 % de las Igs séricas totales y junto a la IgD es la más frecuentemente encontrada en la superficie de los linfocitos B como inmunoglobulina de membrana.
Inmunoglobulina A.
Esta inmunoglobulina posee capacidad neutralizante y precipitante, mientras que su capacidad de fijar complemento y de opsonización son muy débiles, limitándose su efecto a neutrófilos y no a macrófagos.
La propiedad más importante de esta inmunoglobulina viene determinada por su capacidad de unirse por el extremo Fc a la pieza secretora, gracias a la cual puede ser secretada por las mucosas y glándulas exocrinas, ejerciendo su acción más importante en la superficie de mucosas y líquidos biológicos (sobre todo IgA2), tales como el liquido cefaloraquideo, secreción bronquial, lágrima, saliva, etc. Esto es importante porque así protegen precisamente los puntos más vulnerables del organismo, esto es, las puertas de entrada al mismo, como son ojos, boca, aparato digestivo, sistema respiratorio, vagina, etc. No olvidemos que, por ejemplo, si desplegamos la mucosa del aparato respiratorio, la superficie que cubriríamos es de unos 300 m2, superficie que se encuentra en contacto directo con el exterior a través del aire que se respira. Se deduce de ello que, sin duda, deben ser importantes los mecanismos de defensa local entre los cuales la IgA tiene un papel esencial.
Esta inmunoglobulina se encuentra también en la leche materna. Los niveles de todas las inmunoglobulinas, a excepción de la IgG en recién nacidos son muy bajos, siendo por tanto de gran significación el hecho de que la IgA se transfiera desde la madre al lactante a través de la secreción láctea. De ahí que tengamos que insistir en que los lactantes se amamanten en el mayor grado posible directamente por las madres y no con leche de otros orígenes, a lo que actualmente existe excesiva tendencia.
La IgA recibida de la madre ejerce un importante papel de defensa a nivel de todo el aparato digestivo. En ello parece que influyen las especiales características de pH gástrico del lactante que es menos ácido que en el adulto y una especial resistencia de esta inmunoglobulina frente al mismo, por lo que no se destruye a su paso por el estómago.
Inmunoglobulina D.
La concentración de esta inmunoglobulina en suero es muy baja. Hasta fechas muy recientes no se había demostrado que esta inmunoglobulina poseía capacidad de unirse a antígenos, por lo que se dudaba de que actuase con función de anticuerpo. Sin embargo, aunque actualmente se ha demostrado su acción de anticuerpo, no se conoce con precisión cuáles son sus funciones específicas, aunque se piensa que colabora de forma importante en la activación de linfocitos B al actuar como receptor en la superficie de los mismos.
Inmunoglobulina E.
En muchos individuos alérgicos esta inmunoglobulina se presenta en grandes cantidades. El estímulo para su síntesis puede proceder de una gran variedad de antígenos, a los que en este caso se conocen como alergenos. Estos alergenos pueden penetrar en el organismo a través de la piel o de las mucosas respiratoria, ocular, del aparato digestivo, etc., así como por inyectables, como es el caso de la penicilina u otros medicamentos.
La vida media de la IgE en sangre periférica es de 24-48 horas. No tiene capacidad de atravesar la placenta, por lo tanto, las reacciones de hipersensibilidad inmediata no pueden transferirse de manera pasiva de la madre al feto. Sin embargo, puede existir una predisposición de tipo familiar a padecer enfermedades de naturaleza alérgica. Esta predisposición parece estar relacionada con una tendencia a producir anticuerpos de tipo IgE en la respuesta secundaria frente a antígenos, en lugar de IgG que seria la respuesta normal en individuos no alérgicos.
La IgE se encuentra en forma libre en sangre, tambien en otros líquidos biológicos así como unida a basófilos y células cebadas, gracias a la propiedad que tiene esta inmunoglobulina de unirse por su extremo Fc a receptores de superficie presentes en dichas células. Estas células se caracterizan por encontrarse en la piel y mucosas y por contener abundantes gránulos citoplasmáticos, ricos en sustancias vasoactivas que liberan una vez se activan.
dEsnAturAliZaCIóN dE prOtEínaS
Cuando la proteína no ha sufrido ningún cambio en su interacción con el disolvente, se dice que presenta una estructura nativa. Se llama desnaturalización de las proteínas a la pérdida de las estructuras de orden superior (secundaria, terciaria y cuaternaria), quedando la cadena polipeptídica reducida a un polímero estadístico sin ninguna estructura tridimensional fija.
Cualquier factor que modifique la interacción de la proteína con el disolvente disminuirá su estabilidad en disolución y provocará la precipitación. Así, la desaparición total o parcial de la envoltura acuosa, la neutralización de las cargas eléctricas de tipo repulsivo o la ruptura de los puentes de hidrógeno facilitará la agregación intermolecular y provocará la precipitación. La precipitación suele ser consecuencia del fenómeno llamado desnaturalización y se dice entonces que la proteína se encuentra desnaturalizada.
En una proteína cualquiera, la estructura nativa y la desnaturalizada tan sólo tienen en común la estructura primaria, es decir, la secuencia de AA que la componen. Los demás niveles de organización estructural desaparecen en la estructura desnaturalizada.
La desnaturalización provoca diversos efectos en la proteína:
Cualquier factor que modifique la interacción de la proteína con el disolvente disminuirá su estabilidad en disolución y provocará la precipitación. Así, la desaparición total o parcial de la envoltura acuosa, la neutralización de las cargas eléctricas de tipo repulsivo o la ruptura de los puentes de hidrógeno facilitará la agregación intermolecular y provocará la precipitación. La precipitación suele ser consecuencia del fenómeno llamado desnaturalización y se dice entonces que la proteína se encuentra desnaturalizada.
En una proteína cualquiera, la estructura nativa y la desnaturalizada tan sólo tienen en común la estructura primaria, es decir, la secuencia de AA que la componen. Los demás niveles de organización estructural desaparecen en la estructura desnaturalizada.
La desnaturalización provoca diversos efectos en la proteína:
- Cambios en las propiedades hidrodinámicas de la proteína: aumenta la viscosidad y disminuye el coeficiente de difusión.
- Una drástica disminución de su solubilidad, ya que los residuos hidrofóbicos del interior aparecen en la superficie.
- Pérdida de la propiedades biológicas.


Una proteína desnaturalizada cuenta únicamente con su estructura primaria. Por este motivo, en muchos casos, la desnaturalización es reversible ya que es la estructura primaria la que contiene la información necesaria y suficiente para adoptar niveles superiores de estructuración. El proceso mediante el cual la proteína desnaturalizada recupera su estructura nativa se llama renaturalización. Esta propiedad es de gran utilidad durante los procesos de aislamiento y purificación de proteínas, ya que no todas la proteínas reaccionan de igual forma ante un cambio en el medio donde se encuentra disuelta. En algunos casos, la desnaturalización conduce a la pérdida total de la solubilidad, con lo que la proteína precipita. La formación de agregados fuertemente hidrofóbicos impide su renaturalización, y hacen que el proceso sea irreversible.
Los agentes que provocan la desnaturalización de una proteína se llaman agentes desnaturalizantes. Se distinguen agentes físicos (calor) y químicos (detergentes, disolventes orgánicos, pH, fuerza iónica). Como en algunos casos el fenómeno de la desnaturalización es reversible, es posible precipitar proteínas de manera selectiva mediante cambios en:
la polaridad del disolvente
la fuerza iónica
el pH
la temperatura
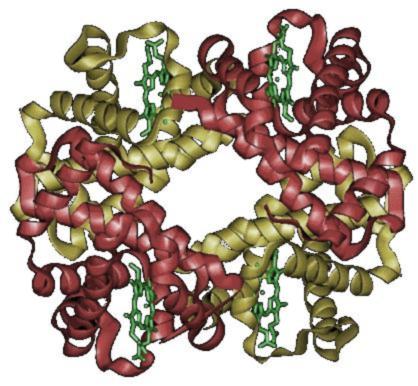
hEmoGLobiNa (Hb)
Es una proteína conjugada de color rojo, es el componente principal de los eritrocitos en un 31 - 34% con un peso molecular de 65000 aprox., sirve de vehículo para el transporte de O2 y CO2.
- Proteína de transporte de Oxígeno que se encuentra en los eritrocitos.
- Pigmento que confiere a la sangre su color rojo.
- Cada eritrocito contiene casi 280 millones de moléculas de hemoglobina.
- Se encuentra disuelta en el citoplasma de los eritrocitos.
- Una molécula de hemoglobina consta de una proteína, GLOBINA; compuesta de cuatro cadenas de polipeptidos (dos cadenas alfa y dos beta) no proteícos, HEM.
- Cada grupo HEM guarda relación con una cadena polipeptídica y contiene un ion Fierro que se combina de manera reversible con una molécula de Oxígeno.
- El oxígeno que se absorbe en los pulmones se trasporta de este modo a los diversos tejidos, en ellos se invierte la unión de del Hierro con el Oxígeno, que se libera de la Hemoglobina y difunde hacia el líquido intersticial y de este a las células.
- También transporta casi 23% del total corporal de CO2 (producto de desecho del metabolismo).
- La sanngre que fluye por los capilares de los tejidos capta el CO2, una parte del cual se combina con aminoácidos de la porción globina de la hemoglobina.
- Al pasar la sangre por los pulmones, el CO2 se separa de la hemoglobjna y se exhala, también participa en la regulación de la presión arterial. Los iones de Hierro de la porción HEM tienen gran afinidad con el Oxido Nítrico (NO), gas que producen las células endoteliales.
- Las concentraciones de hemoglobinas estan influidas por variaciones fisiológicas como la edad, sexo, deshudratación, postura, altitud y por procesos patológicos (anemia y policitemias).
- Se observan valores de hemoglobina elevados en policitemia vera, eritrocitosis, deshidratación, recién nacidos, cianosis congénita o adquirida, enfermedad renal y pulmonar crónica, quistes renales y en una serie de tumores produstores de eritropoyetina.
- Las personas que viven en lugares de de altitud superior a los 1000 m muestran valores significativamente mas elevados.
- En mujeres sanas embarazadas los valores son mas bajos
- Interferencias: la bilirrubina no interfiere, la lipemia puede originar resultados falsamente elevados, debido a la turbidez. Medicamentos y sustancias pueden interferir.
Síntesis:
Inicia en el pronormoblasto (cromoproteína), en la fase del retículocito, responsable del transporte de O2 de los pulmones a los tejidos, mientras que el CO2 va de los tejidos a los pulmones, mediante la circulación sanguínea.
La magnitud de este intercambio gaseoso es directamente proporcional a la concentración de Hb en sangre (en estados patológicos de adición de otros compuestos como el monóxido de carbono, esta regla se rompe) por lo tanto su determinación es un procedimiento para estimar la eficiencia de la circulación sanguínea.
El ferrocianuro de potasio convierte al hierro ferroso de la Hb en hierro ferroso, formando la METAHEMOGLOBINA, que al combinarse con el cianuro de potasio se transforma en CIANOMETAHEMOGLOBINA, es bastante estable y fotocolorimétrico para ser medido su concentración.
La concentración de hemoglobina se calcula a partir de la fórmula:
- Con patrón:
A Muestra X C Patrón
- A patrón
- Sin patrón:
A Muestra X 37.5= C Muestra
Valores de referencia.
Honbres 22.5 a 17.5 g/dl
Mujeres 11.5 a 15.5 g/dl
{kind=link}
Suscribirse a:
Entradas (Atom)